A.J. Rotunno
Mfg./Commissioning/Validation Consultant to the Biopharmaceutical Industry. Past: Biotech Manufacturing: Cell Culture-Three large-scale facility start-ups
@srqstockpicker
Michael Goodman
Michael Goodman
@mikegoodma
Biopharm writer & analyst formerly with Pink Sheet & IN VIVO. Founder/owner of WORD CONTROL - life science communications & market intelligence
http://www.researchgate.net/post/Lentivirus_vs_AAV_for_protein_expression_and_RNAi
serotype
血清分型; 血清型
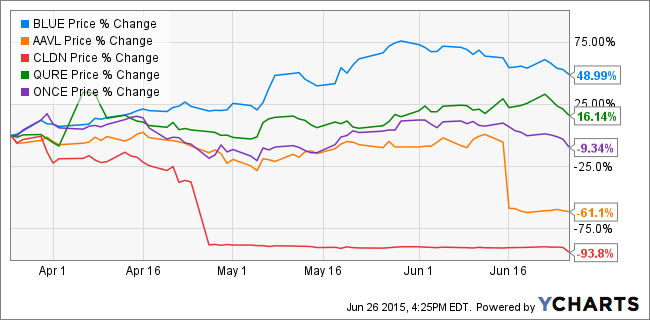
@AlpBugraBasat But that difference is not the reason for the stock outcomes...@SayYayToDrugs@AlpBugraBasat Exactly! If there were multiple -lenti developers then you'll find losers among them too.$QURE best-of-class.@Prof_Dollar@SayYayToDrugs Buy more AAV gene tx companies. This graph will get even worse. :)




Lentivirus 是什麼?
由 Tomislav Mestrovicć, MD, PhD 博士
病毒是小的強制由一件防護蛋白質外套包圍與核糖核酸或脫氧核糖核酸染色體的細胞內寄生生物并且調用他們的基因到被傳染的細胞。
Lentivirus 用長的潛伏期 (甚而幾個月,幾年) 和傾向導致各種各樣的病理學表示慢病毒屬用不同的動物種類。 由於他們的相當靈活的染色體和轉換 nondividing 的細胞, lentiviruses 的許多表單潛在请有成為的一個基因調用的最用途廣泛的向量。
根據國際委員會的最近分類在分類學的病毒 (ICTV), Lentivirus 屬屬於這個系列 Retroviridae 和當前包括九個種類: 七動物 lentiviruses 和二人力 lentiviruses。
動物 lentiviruses 是遲鈍的免疫缺乏病毒 (BIV)、山羊關節炎腦炎病毒 (CAEV)、馬感染貧血症病毒 (EIAV)、似貓的免疫缺乏病毒 (FIV)、美洲獅 lentivirus (PLV)、猿免疫缺陷病毒 (SIV)和 visna/maedi 病毒 (VMV)。 人力種類是著名的人力 immunodeficinecy 病毒 1 (HIV-1) 和 HIV 2 (HIV-2)。
密切相關的人力和猿免疫缺陷病毒幾後裔 (HIV 和 SIV,分別) 在靈長目被找到,能導致獲取的免疫缺乏綜合症狀 (AIDS)。 猿免疫缺陷病毒分支到被命名達成協議他們的主機的五另外的後裔; 那些是非洲青猴、黑猩猩、煤煙灰白眉猴、 syke 和 mandrill SIVs。
靈長目自然感染這些病毒通常保持健康,不用像幫助的疾病的發展,但是他們的對人的傳輸提升了二基因上不同的病毒 (HIV-1 和 HIV-2),密切相關對他們的祖先種類。 這兩病毒在人能導致艾滋病,雖然他們的病毒動能學和流行潛在極大有所不同。
然而,雖然 lentiviruses 對一个負責在人類的歷史記錄的最壞的大流行病,他們為基因治療和基因疫苗的發展廣泛地今天使用。 他們的有用性由這個情況反射他們是無的病毒蛋白質和穩定地合併到靶細胞裡。
此外, lentiviral 向量 (開發從上述複雜逆轉錄酶病毒) 有解決簡單的 retroviral 向量的限制的屬性。 這些包括高穩定性病毒微粒和 insertional 突變形成一個更低的頻率。 這個情況他們可以轉換高度被區分的細胞例如樹狀細胞是關鍵的他們的用於免疫療法。
當輸入這個細胞後,發行一個 lentiviral 核心,并且反向副本進行。 這個核心的基因被運輸對中堅力量,它在稱原病毒綜合化的進程中集成寄主細胞染色體。 副本從集成原病毒,導致與輸入酶和結構上的蛋白質全長核糖核酸染色體的運輸或者通訊員進行 RNAs 形成 (轉換)。
結構上的蛋白質形成了在轉換程序包期間,當 virions 從這個細胞的表面時發芽,因而釋放他們的全長核糖核酸染色體到周圍。 在他們裡面病毒蛋白酶將施加其在特定 polyproteins 的活動,解放結構上的蛋白質和
病毒是小的強制由一件防護蛋白質外套包圍與核糖核酸或脫氧核糖核酸染色體的細胞內寄生生物并且調用他們的基因到被傳染的細胞。
Lentivirus 用長的潛伏期 (甚而幾個月,幾年) 和傾向導致各種各樣的病理學表示慢病毒屬用不同的動物種類。 由於他們的相當靈活的染色體和轉換 nondividing 的細胞, lentiviruses 的許多表單潛在请有成為的一個基因調用的最用途廣泛的向量。
根據國際委員會的最近分類在分類學的病毒 (ICTV), Lentivirus 屬屬於這個系列 Retroviridae 和當前包括九個種類: 七動物 lentiviruses 和二人力 lentiviruses。
動物 lentiviruses 是遲鈍的免疫缺乏病毒 (BIV)、山羊關節炎腦炎病毒 (CAEV)、馬感染貧血症病毒 (EIAV)、似貓的免疫缺乏病毒 (FIV)、美洲獅 lentivirus (PLV)、猿免疫缺陷病毒 (SIV)和 visna/maedi 病毒 (VMV)。 人力種類是著名的人力 immunodeficinecy 病毒 1 (HIV-1) 和 HIV 2 (HIV-2)。
Lentivirus 歷史記錄和特性
在 1843年一 lentiviral 作用者造成的疾病的第一個實際說明來自馬感染貧血症的觀察在法國。 以後,在綿羊的遲緩地累進紊亂在冰島注意在 VMV 造成的 20 世紀 50 年代期間,表示 pneumo 腦病的一份嚴重表單。 在 20 世紀 60 年代,美國研究員確定了白血病牛原因論和 lymphosarcoma - 這個病因因素是逆轉錄酶病毒 (BIV),形態上類似於 VMV。密切相關的人力和猿免疫缺陷病毒幾後裔 (HIV 和 SIV,分別) 在靈長目被找到,能導致獲取的免疫缺乏綜合症狀 (AIDS)。 猿免疫缺陷病毒分支到被命名達成協議他們的主機的五另外的後裔; 那些是非洲青猴、黑猩猩、煤煙灰白眉猴、 syke 和 mandrill SIVs。
靈長目自然感染這些病毒通常保持健康,不用像幫助的疾病的發展,但是他們的對人的傳輸提升了二基因上不同的病毒 (HIV-1 和 HIV-2),密切相關對他們的祖先種類。 這兩病毒在人能導致艾滋病,雖然他們的病毒動能學和流行潛在極大有所不同。
然而,雖然 lentiviruses 對一个負責在人類的歷史記錄的最壞的大流行病,他們為基因治療和基因疫苗的發展廣泛地今天使用。 他們的有用性由這個情況反射他們是無的病毒蛋白質和穩定地合併到靶細胞裡。
此外, lentiviral 向量 (開發從上述複雜逆轉錄酶病毒) 有解決簡單的 retroviral 向量的限制的屬性。 這些包括高穩定性病毒微粒和 insertional 突變形成一個更低的頻率。 這個情況他們可以轉換高度被區分的細胞例如樹狀細胞是關鍵的他們的用於免疫療法。
壽命週期
為了開始其壽命週期,這個感染 lentiviral 微粒 (virion) 必須首先輸入這個細胞。 在病毒束縛到特定感受器官後,那發生通過 endocytosis 或由與細胞膜的直接融合。 對於 HIV-1,感受器官是 T 細胞表面淋巴細胞標記 CD4,雖然病毒也需要 chemocine coreceptors (CXCR4/CCR5)。當輸入這個細胞後,發行一個 lentiviral 核心,并且反向副本進行。 這個核心的基因被運輸對中堅力量,它在稱原病毒綜合化的進程中集成寄主細胞染色體。 副本從集成原病毒,導致與輸入酶和結構上的蛋白質全長核糖核酸染色體的運輸或者通訊員進行 RNAs 形成 (轉換)。
結構上的蛋白質形成了在轉換程序包期間,當 virions 從這個細胞的表面時發芽,因而釋放他們的全長核糖核酸染色體到周圍。 在他們裡面病毒蛋白酶將施加其在特定 polyproteins 的活動,解放結構上的蛋白質和
Popular Answers
Time course of LV expression in vivo and in vitro, two parameters:
- Time until you can detect expression (depends on your detection method)
- Time until expression has reached steady-state level
Pre-expression steps:
Both are slower than the "transient-transfection-of-naked-plasmid" scenario. AAV will not go through the (slow) reverse-transcription step of retro- (including lenti)viruses. But still the ssDNA genome needs to be converted into dsDNA, involving at least second-strand synthesis. Self-complementary AAV vectors (scAAV) have been made to sidestep in-vivo 2nd-strand synthesis, but are suitable only for small constructs (packaging capacity ITR to ITR: 4.7 kb for regular AAV, roughly half this size in scAAV. LTR to LTR in LV: 10.2 kb)
Integrated LV:
The single most influential factor on the onset of LV expression is the strength of your internal promoter, in the target cell type. For instance in neurons, lentiviral GFP driven by Synapsin promoter will be visible (using ordinary epifluorescence) ca. 30 h after transduction, steady state will be reached in ca. 48 h. GFP driven by CAG promoter, visible ca. 48 h, steady state ca. 4 days. So the rate of onset may be very different in two vectors of the same type (LV), depending on transcriptional efficiency.
AAV:
Serotypes differ dramatically in their ability to target various tissues and cell types. Choosing the right serotype for your target tissue will make the vector phenotypically "faster" because transduction efficiency is higher.
Spread:
Basically depends on two things, particle size and tissue density. Particle sizes: rAAV, 20 nm; LV, >100 nm. Smaller AAV particles will spread further (in a given tissue) than larger LV particles. Tissue density depends on brain region (e.g. directional spread in white matter), but most of all, on age. In embryonic to early-postnatal brains, LV will spread a lot (and AAV likewise, I suppose). In older animals, the difference may become more noticeable. In the adult, LV practically no longer spreads at all.
Duration:
In postmitotic cells, AAV expression is fairly stable. Colleagues have repeatedly imaged the same AAV-transduced cortical neurons in vivo, for weeks. In dividing cells, the AAV genome will not replicate along with the host genome, and expression will get lost depending on cellular growth rate (faster in fast-dividing cells).
I'm afraid there is no Jack-of-all-trades vector ;)