一般的观点是说变异的产生是随机的,自然选择决定进化方向。今天看了一篇博文 http://www.geekonomics10000.com/591 提到了一个新的观点「基因变异有特定的方向」,靠谱么?
传统教科书中的自然选择,说基因突变完全随机,进化是为适应环境来决定哪种变异被保留。而在过去三十年,科学家开始使用非线性数学和计算机模拟的手段来研究进化论,其得出的最关键思想,就是进化不是完全随机的。所有动物的视网膜上都有同一种叫做视紫红质的特殊蛋白质,它的作用是把眼睛接收到的光能变成电信号传输给视觉神经。在所有可能处理光信号的蛋白质分子中,视紫红质的性能是最好的,生物进化早在几十亿年前就发现了这个完美分子的结构,而且从没有变过。如果进化是完全随机的,那么在所有可能的蛋白质分子中找到这么一个完美分子,就好像在茫茫宇宙中找到一颗特定恒星一样困难。这还不是最可怕的。分子生物学的研究表明,视紫红质是在古细菌和真细菌这两个进化路线上完全独立的分支上分别被进化出来的。也就是说进化不但找到了这个分子,而且还找到了两次!从统计角度完全随机的进化绝对做不到这一点。
所以有些最新的进化论学说认为生物通过细胞的新陈代谢之类的过程,存在一个自组织的机制,使得基因变异有一个特定的方向。而这种学说的关键证据,在于生命组织的形成方法是有限的。
按投票排序按时间排序
18 个回答

有意思,这博客里的很多故事我都知道,我也对历史的必然和偶然感兴趣,但是,同样的例子,我的结论不尽相同。
先说一个,
而上文中这三种翅膀,还正好是脊椎动物中三个不同的翅膀进化方式:蝙蝠是手成翅,鸟是胳膊成翅,而翼手龙是指头成翅。至于这算不算是一样结构,见仁见智。
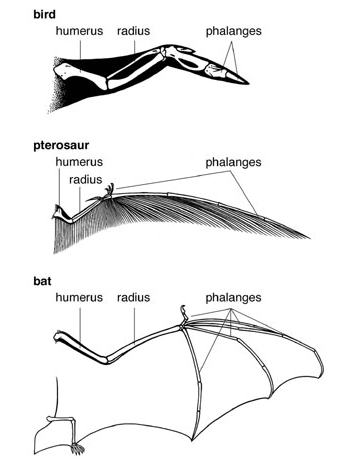
(图片取自 Sean Carroll 的 Endless Forms Most Beautiful)
再比如:
对应理论是,要么这个 Pax6,或者说整个 Pax 家族,被不同物种反复发现,要么就是大家都是从一个根出来的。
我倾向于后一种。不能说全中,但绝大部分情况下,这些重要的基因单元,都是同源的。
再比如,文中说:
人:RHO rhodopsin [ Homo sapiens ]
http://www.ncbi.nlm.nih.gov/sites/entrez?db=gene&cmd=retrieve&dopt=default&rn=1&list_uids=6010
小鼠:Rho rhodopsin [ Mus musculus ]
http://www.ncbi.nlm.nih.gov/sites/entrez?db=gene&cmd=retrieve&dopt=default&rn=1&list_uids=212541
用 blastn 作基因比对,序列很长,结果我只取第一段 540 个核酸:

可以看见,相似度极高,83%,p-value 是 10-152,说明是同源的,但,怎么也不能说是从没变过吧?只能说是高度保守,功能极其相似。这不恰恰是进化选择的结果么。
如果不是这样的话,所有的基因序列分析,什么同源基因理论,都没法玩了。当然不是说这些既有的理论一定对,而是说,这文中的理论,如果只按这字面的理解,绝不是进化生物的主流。
事实上,按这理论,我完全就可以导出神创论出来:既然进化不随机,那为什么偏偏是最完美的结果被选中呢?那一定是幂幂中有人定的。
你可以去信这个,但不要跟进化论混在一起。
最后说,变异有没有方向。
当然有的。
比如 ACGT 四个核酸,共价键的强弱都不一样,加上前后核酸的影响,当然发生变异的机会是不是样的。所谓随机总有个分布,而分布并不是想像中的,所有可能都一样,而方向,也不是说一定会怎么样,而只是某种可能更大一点。像紫外线照射,就会制造著名的 “UV signature” 变异,就是指 C->T 变异,而少有其他变异,这也是很多黑色素瘤的特征。
当然,这个只是在变异尺度上的,这个方向,和我们通常说的在功能尺度上的方向,是有相当的距离的,这样看,说这些变异本身没有特定的方向,也没问题。
最终,变异和选择是一个复杂的体系,变异的方向性,还是要同时看生存压力,是否有进化优势。变异只是个起点。
随手写写,有很多不严谨的地方,大家见谅。 显示全部
先说一个,
再比如说翅膀,世界上可能只有一种形成翅膀的方法,所以蝙蝠、鸟类和翼手龙虽然独立进化,其翅膀结构却是一样的。这里的翅膀,显然只是脊椎动物的。
而上文中这三种翅膀,还正好是脊椎动物中三个不同的翅膀进化方式:蝙蝠是手成翅,鸟是胳膊成翅,而翼手龙是指头成翅。至于这算不算是一样结构,见仁见智。
(图片取自 Sean Carroll 的 Endless Forms Most Beautiful)
再比如:
人眼这个结构不但出现在哺乳动物中,而且出现在六种不同的生物种类中 — 这六个物种的共同祖先是没有眼睛的,它们是在进化史上分道扬镳以后才各自独立地进化出来了眼睛,而且是同一种眼睛。更进一步,组成眼睛一共就只有九种方法,而这九种方法都被进化所发现了。这眼有多少种结构,我是不知道的,但我知道,这眼睛自打五亿年前出现,到现在,决定眼睛长在哪里的基因,就是 Pax6,不知道是不是所有动物,但应该基本都是如此。
对应理论是,要么这个 Pax6,或者说整个 Pax 家族,被不同物种反复发现,要么就是大家都是从一个根出来的。
我倾向于后一种。不能说全中,但绝大部分情况下,这些重要的基因单元,都是同源的。
再比如,文中说:
在所有可能处理光信号的蛋白质分子中,视紫红质的性能是最好的,生物进化早在几十亿年前就发现了这个完美分子的结构,而且从没有变过。好,那我取两个视紫红质基因序列,就是人的和小鼠的:
人:RHO rhodopsin [ Homo sapiens ]
http://www.ncbi.nlm.nih.gov/sites/entrez?db=gene&cmd=retrieve&dopt=default&rn=1&list_uids=6010
小鼠:Rho rhodopsin [ Mus musculus ]
http://www.ncbi.nlm.nih.gov/sites/entrez?db=gene&cmd=retrieve&dopt=default&rn=1&list_uids=212541
用 blastn 作基因比对,序列很长,结果我只取第一段 540 个核酸:
可以看见,相似度极高,83%,p-value 是 10-152,说明是同源的,但,怎么也不能说是从没变过吧?只能说是高度保守,功能极其相似。这不恰恰是进化选择的结果么。
如果不是这样的话,所有的基因序列分析,什么同源基因理论,都没法玩了。当然不是说这些既有的理论一定对,而是说,这文中的理论,如果只按这字面的理解,绝不是进化生物的主流。
事实上,按这理论,我完全就可以导出神创论出来:既然进化不随机,那为什么偏偏是最完美的结果被选中呢?那一定是幂幂中有人定的。
你可以去信这个,但不要跟进化论混在一起。
最后说,变异有没有方向。
当然有的。
比如 ACGT 四个核酸,共价键的强弱都不一样,加上前后核酸的影响,当然发生变异的机会是不是样的。所谓随机总有个分布,而分布并不是想像中的,所有可能都一样,而方向,也不是说一定会怎么样,而只是某种可能更大一点。像紫外线照射,就会制造著名的 “UV signature” 变异,就是指 C->T 变异,而少有其他变异,这也是很多黑色素瘤的特征。
当然,这个只是在变异尺度上的,这个方向,和我们通常说的在功能尺度上的方向,是有相当的距离的,这样看,说这些变异本身没有特定的方向,也没问题。
最终,变异和选择是一个复杂的体系,变异的方向性,还是要同时看生存压力,是否有进化优势。变异只是个起点。
随手写写,有很多不严谨的地方,大家见谅。 显示全部

进化不是随机的,但突变是随机的。
在你的问题中,这两个概念被严重混淆了。
你所说的“突变不是随机的”,其中的“突变”指的不是历史上发生过的全部突变,而是那些已经被自然选择固定下来的有利突变。
在这个意义上来说,“突变”的确不是随机的,因为自然选择不是随机的。
这种非随机性体现在进化的结果上是很明显的。视紫红质是一个例子,但不像你想的那么惊人。蛋白质分子中殊途同归的例子远不止这一个。
在宏观性状上,这样的例子也很多,称为趋同进化。比如飞鼠的翼膜、鼹鼠的眼睛,都是高度特化的器官,但在澳大利亚的有袋类哺乳动物中,都独立进化出了同样会飞的翼膜和退化的眼睛。这样的性状牵涉到更多的基因和蛋白的改变,在我看来比视紫红质的趋同进化更“不可思议”。
多说一句,这种自然选择的非随机性体现为进化的有向性。注意,不是从低级到高级这种有向性(这在进化论中是错误的概念),而是向着更适应环境需求的方向进化。这里的”环境需求“是广义的,比如在视紫红质的例子上,就是指对于高效感应光线的需求。
这种有向性正是我主张evolution应翻译为“进化论”的主要原因。(参见我对另一个问题的回答:http://www.zhihu.com/question/20214570/answer/14360326)
有人总结说,进化论是“对随机突变的非随机选择”。我想这句话是对这个问题的最好回答。
在你的问题中,这两个概念被严重混淆了。
你所说的“突变不是随机的”,其中的“突变”指的不是历史上发生过的全部突变,而是那些已经被自然选择固定下来的有利突变。
在这个意义上来说,“突变”的确不是随机的,因为自然选择不是随机的。
这种非随机性体现在进化的结果上是很明显的。视紫红质是一个例子,但不像你想的那么惊人。蛋白质分子中殊途同归的例子远不止这一个。
在宏观性状上,这样的例子也很多,称为趋同进化。比如飞鼠的翼膜、鼹鼠的眼睛,都是高度特化的器官,但在澳大利亚的有袋类哺乳动物中,都独立进化出了同样会飞的翼膜和退化的眼睛。这样的性状牵涉到更多的基因和蛋白的改变,在我看来比视紫红质的趋同进化更“不可思议”。
多说一句,这种自然选择的非随机性体现为进化的有向性。注意,不是从低级到高级这种有向性(这在进化论中是错误的概念),而是向着更适应环境需求的方向进化。这里的”环境需求“是广义的,比如在视紫红质的例子上,就是指对于高效感应光线的需求。
这种有向性正是我主张evolution应翻译为“进化论”的主要原因。(参见我对另一个问题的回答:http://www.zhihu.com/question/20214570/answer/14360326)
有人总结说,进化论是“对随机突变的非随机选择”。我想这句话是对这个问题的最好回答。

从外在来说,存在选择的压力,所以理论上是随机事件的基因突变,会被选择所修正;从内因上说,某些片段和序列倾向于发生突变(比如存在回文结构,链内配对,以及生理条件下跟蛋白相互作用时的构象变化等等),或者是某一个基因突变后,增加了整个基因组的不稳定性,这都是所谓“定向性突变”的基础。
假设引文中没有虚假信息
回答提问者问题:
DNA碱基对的突变是随机的,新性状的出现已经有了一定的方向(致死突变都死在娘胎了),进化的方向是由生存压力和中性漂变决定的。
而在过去三十年,科学家开始使用非线性数学和计算机模拟的手段来研究进化论,其得出的最关键思想,就是进化不是完全随机的。这句话说的是“进化”不是完全随机的,起先我想,这个我们知道很久了,过去三十年得到了什么东西?然后看到“非线性数学和计算机模拟的手段“,猜测是指中性漂变,可以理解为没有生存压力的前提下,进化出现方向性的概率仍然不小。
生物进化早在几十亿年前就发现了这个完美分子的结构,而且从没有变过。这句话说的不就是选择压力使得这个分子维持现状咩?
如果进化是完全随机的,那么在所有可能的蛋白质分子中找到这么一个完美分子,就好像在茫茫宇宙中找到一颗特定恒星一样困难。这还不是最可怕的。分子生物学 的研究表明,视紫红质是在古细菌和真细菌这两个进化路线上完全独立的分支上分别被进化出来的。也就是说进化不但找到了这个分子,而且还找到了两次!从统计 角度完全随机的进化绝对做不到这一点。这个观点和所有“进化出智能生物的概率太小所以一定有神创”的观点一样,概率算错了。
所以有些最新的进化论学说认为生物通过细胞的新陈代谢之类的过程,存在一个自组织的机制,使得基因变异有一个特定的方向。这句话前言不搭后语,“自组织”现在就是一个筐,什么都往里丢,都不去搞搞明白什么是自组织。这也不是什么新概念了,前面我说概率算错了,也是因为计算的时候没有考虑实际过程(“自组织”以及优化出来的东东会被保留下来)。
而这种学说的关键证据,在于生命组织的形成方法是有限的。这句话真心没看懂。
回答提问者问题:
DNA碱基对的突变是随机的,新性状的出现已经有了一定的方向(致死突变都死在娘胎了),进化的方向是由生存压力和中性漂变决定的。



如果眼睛在突变中产生非“视紫红质”的另外一种物质。比如“视青红质”吧。
那么在生存斗争中,“视青红质”的效率是不如“视紫红质”的。这就给携带“视青红质”的生物带来生存压力。
死掉了,就没法传承了。。
几十代下来,就只剩“视紫红质”了
那么在生存斗争中,“视青红质”的效率是不如“视紫红质”的。这就给携带“视青红质”的生物带来生存压力。
死掉了,就没法传承了。。
几十代下来,就只剩“视紫红质”了

同一个个体内,不同的视紫红质单体的结构肯定也有差别。
结构相似就能达成功能,并不是在数轴上随机出一个精确的点来,因此凭啥不能被进化出来?何况,选择出这玩意儿的概率并不是均等的,基因确实可以突变出别的分子,但别的分子活下来的可能又有多少?
如果在不同的计算机内用穷举法找出符合当前需求、性能最佳的结果,相信结果也是大体相同的,不论你在多少计算机上做实验。
穷举最可怕的地方就是大浪淘沙,绝不会错过一个。
结构相似就能达成功能,并不是在数轴上随机出一个精确的点来,因此凭啥不能被进化出来?何况,选择出这玩意儿的概率并不是均等的,基因确实可以突变出别的分子,但别的分子活下来的可能又有多少?
如果在不同的计算机内用穷举法找出符合当前需求、性能最佳的结果,相信结果也是大体相同的,不论你在多少计算机上做实验。
穷举最可怕的地方就是大浪淘沙,绝不会错过一个。


基因变异是随机的,而生物进化是有特定方向的。
就比如我们身边的人就有很多因为基因突变而形成的各类 症状,如 侏儒症,巨人症。这些都是不好 的突变,导致不能适应社会, 就会被淘汰,而那些能适应自然环境的基因突变更容易保存下来,通过漫长时间的演变 导致看起来基因突变是有方向的。
就比如我们身边的人就有很多因为基因突变而形成的各类 症状,如 侏儒症,巨人症。这些都是不好 的突变,导致不能适应社会, 就会被淘汰,而那些能适应自然环境的基因突变更容易保存下来,通过漫长时间的演变 导致看起来基因突变是有方向的。

首先我觉得进化本身就不是随机的。因为物种是受到生存选择的压力的,所谓适者生存,不适合的要被淘汰。所以进化是有规律可循的。
另一方面,我倾向于单个碱基的突变是完全随机的,没有任何规律的。这个就是所谓的中性理论的核心吧。注意是碱基突变而不是进化。
但是为什么我们观测的结果是某些碱基特别容易突变或者缺失,我觉得是因为自然选择造成的。可以假设有一个基因,它转录的蛋白控制着生命体的关键性状,如果这个基因某一个关键位置的碱基发生了突变,导致这个基因的表达出现了问题,于是这个生命体就死了。于是这个突变体也就没有将自身传给下一代的可能,也就是说,不存在了。当然大部分时候没有我描述的那么极端。但是可以肯定的是,在真基因中,绝大部分的突变都是有害的,并且能够被保留的突变一般都出现在密码子的第三位碱基,即同意突变。偶然有一些碱基的突变导致生命体对环境适应能力增强,因此该基因被复制遗传给下一代的可能性会大大增加,于是导致了进化。
这是我对中性理论的一点理解。
至于第二个找到两次的问题,我觉得在概率上也是有可能的。DNA复制所需要的酶的错误概率大概在十的七次方分之一这个水平。而人类基因组本身大小在30亿。也就是说我们自身遗传物质复制一次的过程都有可能出现不止一次错误。但是万幸我们有修复和识别错配的机制。但是在一些低等的生物比如细菌中,它的修复机制很不完善,甚至在极端条件下会采取极端措施,保留基因组完整但倾向错误配对,所谓的SOS 修复或者称错误倾向修复。在生命发生的35亿年里,不同的基因得到类似功能的可能性,应该是存在的,并且很可能不想我们想象的那么低。
另一方面,我倾向于单个碱基的突变是完全随机的,没有任何规律的。这个就是所谓的中性理论的核心吧。注意是碱基突变而不是进化。
但是为什么我们观测的结果是某些碱基特别容易突变或者缺失,我觉得是因为自然选择造成的。可以假设有一个基因,它转录的蛋白控制着生命体的关键性状,如果这个基因某一个关键位置的碱基发生了突变,导致这个基因的表达出现了问题,于是这个生命体就死了。于是这个突变体也就没有将自身传给下一代的可能,也就是说,不存在了。当然大部分时候没有我描述的那么极端。但是可以肯定的是,在真基因中,绝大部分的突变都是有害的,并且能够被保留的突变一般都出现在密码子的第三位碱基,即同意突变。偶然有一些碱基的突变导致生命体对环境适应能力增强,因此该基因被复制遗传给下一代的可能性会大大增加,于是导致了进化。
这是我对中性理论的一点理解。
至于第二个找到两次的问题,我觉得在概率上也是有可能的。DNA复制所需要的酶的错误概率大概在十的七次方分之一这个水平。而人类基因组本身大小在30亿。也就是说我们自身遗传物质复制一次的过程都有可能出现不止一次错误。但是万幸我们有修复和识别错配的机制。但是在一些低等的生物比如细菌中,它的修复机制很不完善,甚至在极端条件下会采取极端措施,保留基因组完整但倾向错误配对,所谓的SOS 修复或者称错误倾向修复。在生命发生的35亿年里,不同的基因得到类似功能的可能性,应该是存在的,并且很可能不想我们想象的那么低。

No comments:
Post a Comment