li 理想气体分子只有动能,没势能,当然也全是机械能。因为方向和大小无规,才称为内能
蛋白质传递能量的机制
蛋白质传递能量的机制
在分子凝聚态中,能量的转移并不一定需要依靠直接的电子迁移,只需要从电子激发态跳回到基态,并把释放出来的能量传递给相邻的原子,使后者电子从基态跃迁到激发态,从而实现能量的转移。这种转移表现为电子激发态的迁移。接着第二个原子跳回到基态,把释放的能量传递给第三个原子,使它的电子激发起来……于是形成了一幅电子激发态迁移的图画。这种运动形式叫激子。但是激子的运动是不稳定的,激发能会很快地变成热而耗散掉,亦即靠这种方式迁移电子激发态,其效率往往不高,无法与以激发能耗散为热的过程相匹敌。为使电子激发状态能稳定地传播,必须和分子的构象变化联系起来,依靠构象变化使激子的运动稳定化。这种运动形式叫孤子或孤波。自从1976年达维多夫提出蛋白质中的孤子(孤波)以来,这方面理论工作进行了很多,但还没有确切的实验。达维多夫孤子是构象变化和电子激发的耦合结果。多分子系统中,这种电子跃迁的构象运动集体模式可称为构象子,例如膜磷脂系统中有可能观察到构象子。在链式分子中的能量传输,由于序列是非均匀的,构象振动一般只能局域于某一位点邻近,可把它们看作局域声子。但由于相邻位点的电偶极作用,它们也可传递。这种局域声子激发沿链传输,提供了一种以一系列循序衔接的构象变化为特征的能量转移机制。和构象变化耦合在一起的各种电子激发的运动,是电子在生物体内扮演的重要角色。
弱力是生命活动的基本力,弱键是活性能的贮藏所。因为弱键是容易变化的,生物体的平均热能就可以改变它;而强键(共价键)则是稳固的、不易变化的。但这决不是说共价键在生命活动中不重要。新陈代谢的每一步总伴随着一些强键的破裂和另一些强键的生成。酶的本质就在于它能加速生成或裂解一个特异的共价键。由于酶的催化,那些在一般条件下根本不可能实现的反应都能在生物体内高效率地进行。酶的催化功能和两个因素有关,一是分子构象,二是电子运动,因为被催化的反应总和电子运动有关。
超导基态应该是个电子对形成的相干态。这大概就是灵感。
相干态是个很神奇的量子态,粒子数不定,但具有最小的位相涨落。很多玻色子凝聚相都处于相干态,比如激光
DOC檔
psroc.phys.ntu.edu.tw/bimonth/download.php?d=2&cpid=83&did...
由 郭華丞 著作 - 相關文章
此相變的臨界磁場,和電流電壓特性曲線中所觀察到庫柏電對的庫倫阻斷 及 約 ..... 表示庫伯電子對的傳導是熱激發的,其中 E b 代表一障壁能量,實驗上約為120 ..... 低電阻代表磁通的運動是受到侷限的 , 此狀態稱作磁通的commensurate state[6,7] 。超导体_王为民科学与艺术网站
Photon 2Dphonon Coupling and Photonic Cooper Pairs是什么意思_ ...
phymath999: bcs01 凝聚体中的库伯对占据了费米面以下所有的动量 ...
phymath999.blogspot.com/2014/01/bcs01.html - 轉為繁體網頁
《哈比人》惡龍史矛革大有來頭,班奈狄克庫柏拜區曾經調教娜姐| 星聞 ...
tw.omg.yahoo.com/.../哈比人-惡龍史矛革大有來頭-班奈狄克庫柏拜區...
超導理論 - 新華網搜索
big5.xinhuanet.com/gate/big5/search.news.cn/inSearch.jspa;...?n...
| 3 . 3 生物超导体 | 加到收藏夹 添加相关资源 |
3.3 生物超导体
在一个很长时间内,超导电现象一直令人迷惑不解。事实上,它是一种宏观量子现象,只有依据量子力学才能给出正确的微观解释。1956年,库柏(Cooper)证明:在费米面上的一对电子,如果它们之间存在净吸引力,不管多么微弱,这对电子将形成一束缚态。他从理论上进一步指出,两个具有大小相等、方向相反的动量和自旋的电子间存在着最强的吸引力。1957年,巴丁(Bardeen)、库柏(Cooper)和徐瑞弗(Schrieffer)成功地解决了超导电机制,建立了第一个超导电性微观理论——BCS理论,共同获得1972年诺贝尔物理奖。
生物体内存在着超导体吗?一直是个引人入胜的问题。1964年,勒特耳(Little)基于伦登(Lon-don)提出的超导态有可能在大的有机分子如蛋白质中存在的思想,首先设想由一条主链和一系列键合在主链上的侧键组成的有机分子,可能发生激子机制超导电性,而且其临界温度Tc可能很高。
蛋白质分子是由20多种氨基酸连接而成的共价多肽链。其连接的基本方式是肽(图3.1)。蛋白质的满带(最高被电子占据带)和最低带间的能隙约3电子伏以上,比一般半导体大好几倍。蛋白质不可能依靠热运动把电子从满带跳到上面的空带中去,因为缺少可以导电的载流子。所以蛋白质不是半导体而是电介质,即平常所说的绝缘体。
蛋白质传递能量的机制
在分子凝聚态中,能量的转移并不一定需要依靠直接的电子迁移,只需要从电子激发态跳回到基态,并把释放出来的能量传递给相邻的原子,使后者电子从基态跃迁到激发态,从而实现能量的转移。这种转移表现为电子激发态的迁移。接着第二个原子跳回到基态,把释放的能量传递给第三个原子,使它的电子激发起来……于是形成了一幅电子激发态迁移的图画。这种运动形式叫激子。但是激子的运动是不稳定的,激发能会很快地变成热而耗散掉,亦即靠这种方式迁移电子激发态,其效率往往不高,无法与以激发能耗散为热的过程相匹敌。为使电子激发状态能稳定地传播,必须和分子的构象变化联系起来,依靠构象变化使激子的运动稳定化。这种运动形式叫孤子或孤波。自从1976年达维多夫提出蛋白质中的孤子(孤波)以来,这方面理论工作进行了很多,但还没有确切的实验。达维多夫孤子是构象变化和电子激发的耦合结果。多分子系统中,这种电子跃迁的构象运动集体模式可称为构象子,例如膜磷脂系统中有可能观察到构象子。在链式分子中的能量传输,由于序列是非均匀的,构象振动一般只能局域于某一位点邻近,可把它们看作局域声子。但由于相邻位点的电偶极作用,它们也可传递。这种局域声子激发沿链传输,提供了一种以一系列循序衔接的构象变化为特征的能量转移机制。和构象变化耦合在一起的各种电子激发的运动,是电子在生物体内扮演的重要角色。
弱力是生命活动的基本力,弱键是活性能的贮藏所。因为弱键是容易变化的,生物体的平均热能就可以改变它;而强键(共价键)则是稳固的、不易变化的。但这决不是说共价键在生命活动中不重要。新陈代谢的每一步总伴随着一些强键的破裂和另一些强键的生成。酶的本质就在于它能加速生成或裂解一个特异的共价键。由于酶的催化,那些在一般条件下根本不可能实现的反应都能在生物体内高效率地进行。酶的催化功能和两个因素有关,一是分子构象,二是电子运动,因为被催化的反应总和电子运动有关。
激子超导电性
大家知道,电子之间本有库仑排斥力,但在超导体内的两电子间由于交换声子而产生了吸引作用。当这种吸引作用超过两电子间的库仑排斥作用时,两电子就形成电子对引起超导电性,这就是电声子机构的超导电性。1964年,勒特耳首先提出,不是利用交换声子(离子晶格波的能量激发)使两电子间产生吸引,而是使两电子间交换激子而产生吸引作用。勒特耳设想了一种结构的有机分子,它由两部分组成。如图3.9表示长链部分A(或形象地称为“脊椎骨”)是电子导电的主体部分;另一部分是联到“脊椎骨”的一系列旁链B(称为臂)。

假设在旁链分子中有正电荷从B链的左端到右端发生振荡(电荷极化),按照量子力学,这种电荷振荡系统的能量是量子化的。设其基态与第一激发态之间的能量间隔(能量子)为hω,则称hω为激子。在长链A中,电子与产生电荷极化的旁链分子发生相互作用,这是电子-激子相互作用,而通过激子作为中间媒介,可能在两电子之间产生一种净吸引作用。图3.10是两电子通过激子耦合的示意图,图中粗线代表激子系统,|i>表示其基态,|a>表示其激发态。图中左半表示“脊椎骨”中第一个电子与激子系统相互作用,电子1放出一个激子使激子系统从基态|i>激发到|a>态;图中右半部分表示激子系统与电子2相互作用后从激发态|a>回到基态|i>,这时激子被电子2所吸收。这样,电子1和电子2由于交换激子产生一种间接相互作用。如果这种激子机制能产生两电子间的净吸引,那么,可以预期将出现超导态,这就是激子超导电性。

勒特耳对他提出的如图3.11所示的长链和旁链分子作了粗糙的理论估计,结果表明可以产生净的吸引作用,相应的激子超导转变温度Tc可达220K以上!
1971年克柏报道,具有高浓度胆固醇的神经纤维的某些部分,在生理温度下有超导性。后有人进一步报道,实验证实胆酸、脱氧胆酸、石胆酸、胆烷酸钠盐的抗磁性分别在30K、60K、130K、277K起突然变化,突变时原子晶体结构没有变化,本质上应是由电子引起的。氨基酸常温下不导电,如果有的氨基酸的Tc在250K左右,那就表示存在着高温超导区,即材料整体是绝缘体,但材料本体内,分散着小的超导区,称为零星超导体。还有人认为,就脱氧核糖核酸(DNA)的分子结构而言,可能产生所谓的激子超导电性。勒特耳的思想在许多物理和化学工作者中引起了广泛兴趣。在勒特耳设想中,超导电性在“脊椎骨”链中产生。这是一种低维(低于三维)超导体,它和通常讨论的三维晶格超导体很不一样。但是尽管理论上作了很多设想,迄今为止还没有实验事实能够肯定激子超导机制。

| BCS之美(五):为什么没电阻
|||
现在关于BCS超导的争论,已经基本熄火,主要原因是缺氧,因为龚博主不出面。BCS理论看起来很简单,非常直观地解释了低温超导的物理机制,而且BCS对高温超导也有重要的借鉴意义,因为配对机制仍然是基础。这里我讲讲BCS怎么解释超导体的0电阻。
超导体内的电子因为库伯配对,形成一个凝聚体,产生了一个凝聚能。凝聚能可以这样理解,如果你想拆开这些配对,就要最少消耗凝聚能这么多能量。任何一种结合,都有这样一个能量的,比如H原子的一个质子和一个电子结合在一起,有个结合能13.6eV, 如果要拆开一个氢原子,也就是把它的电子敲出去,就要最少消耗13.6eV的能量。超导体中的电流要产生电阻,电能转化热,能量只能来自于凝聚体的动能。如果凝聚体的动能小于刚才讲的凝聚能,凝聚体就没法损失能量,从而产生不了热,于是没电阻。当电流密度足够大,也就是库伯对的漂移速度足够大, 有超导电流的时候,整个凝聚体作为一个整体往前跑。这件事情可以反过来看问题。以凝聚体质心为参考系,晶格所有的正离子往后跑,而且速度一样。 而凝聚体还是原来那个没有电流时候的凝聚体。以晶格为参照系,情况有点复杂,因为凝聚体中的库伯对占据了费米面以下所有的动量值,有各个方向的动量。这个图像让黄秀清很纠结。其实,一个普通材料中电子的动量分布也是有各向均衡的,否则大量电子就有定向的集体运动。黄猴想不通,一个库伯对,动量相反,也就是速度相反,怎么能一起往前跑。其实,一个库伯对,并不是固定的两个电子配成的对,是大量电子的集体行为,电子不断高频碰撞,动量在随时改变,相反的动量总是存在的。 黄猴还质疑电子吸收声子形成库伯对的机制,他觉得这种声子媒介不需要时间的事情不可信。这样一种图像是现代物理学对相互作用的普遍描述,是量子场论的基本内容。比如电磁作用在量子电动力学中表现为电荷之间的光子交换,强作用在量子色动力学中是夸克之间的胶子交换,弱作用在弱电统一理论中是Z,W+,W-这些中间玻色子交换,都是不用时间的。这种描述已经取得了巨大的成就,早已成为现代物理学的基本内容。这些中间交换的媒介粒子的确是虚粒子,不满足质壳关系,所以,是不可以用仪器测量到的。它们只是一种数学描述,称为费曼图。但独立的媒介粒子可以测量到。 BCS之美(四):库伯对
|||
由于电子空穴组合,超导体内部电子在能量上的分布发生了变化。对于非超导材料来讲,温度为0的时候,电子填满在费米能级以下的所有能级。此时,如下图虚线所示,费米能级以上能级是空的,而费米能级以下是满的。但是,如果存在超导凝聚,在费米能级 EF附近,电子分布有一点点改变。此时,如下图红色实线所示,费米能级以上有了电子,费米能级以下有了空穴。
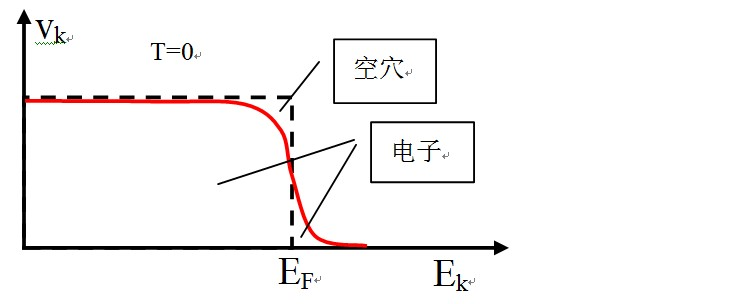
就是这样一个电子分布改变,导致超导体内电子发生凝聚。费米面附近的电子,也就是上图中红色曲线跨过EF附近的电子发生配对,形成动量为0的库伯对。由于它们的动量都是0,这就相当于玻色-爱因斯坦凝聚BEC。
库伯对是很奇特的。一个库伯对中的两个电子并不像氢原子中的电子和质子结合在一起,而是像“潜水”一样,时而一起浮出“水面”时而一起潜入“水下”。一个库伯对可以写成
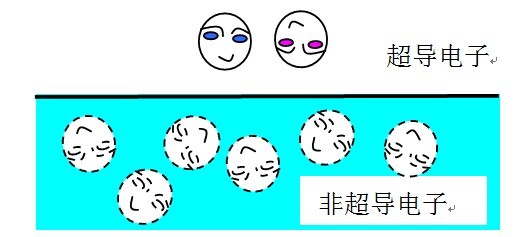
如上图所示,库伯对中的两个电子,就像图中两哥们,在“水面”上下蹦来蹦去,而且他们步调一致。但在“水面”以下,它们就跟其他电子混在一起乱了。可见,这种配对的图像,跟很多人想象的配对是完全不同的。
超导凝聚的关键还在于,所有库伯对与共同的位相,也就是
(注:我写这个系列是个整理思路的过程。我发现,一边写还能一边有一些新想法。这是科普的好处。)
BCS之美(三)
|
No comments:
Post a Comment